『珍藏版』Science綜述|睡眠大腦的記憶調控
來源: BioArt
撰文|姜紅
責編 |酶美
睡眠是人類和動物王國的基礎生理活動。我們的身體可以是處於靜止的、不動的,但是大腦卻是可以十分活躍。哲學家把大腦的狀態分成四個階段:清醒(Awakening),睡眠(Sleep),沉睡(Slumber)和最高境界(Supreme)。睡眠是有多重要呢,文藝復興時期著名的戲劇家和作家威廉莎士比亞就有經典的美妙地關於睡眠的臺詞:

我們爲什麼每天需要躺下,失去意識幾個小時進行休整?睡眠的作用到底是什麼?如果不睡會造成什麼影響?近十年來,得益於技術手段的進步,在理解睡眠和解決睡眠障礙等諸多睡眠大腦相關的問題上都有了很多突破,包括從微觀的分子機制,到宏觀的網絡層次,使我們越來越靠近理解睡眠的真相。在現代社會,睡眠對身體和精神衛生健康的重要性往往是被低估的(996工作機制的危險性問題?):實驗動物學的研究也表明睡眠不足導致認知能力下降; 在一項美國社區研究中所記錄的大量睡眠剝奪的案例,平均睡眠時間只有6.1個小時,而專家建議成人睡眠時間爲7到9個小時。在發展中國家和發達國家的貧困人口中,睡眠的平均時間和睡眠質量也是低的。這就表明,睡眠問題不僅僅 是關係生理健康的問題,也是關係到經濟發展和社會民生問題。
2021年底,Science專欄將焦點聚焦在睡眠的研究上,睡眠領域專家撰寫的最新關於睡眠與學習記憶、鎮靜以及睡眠的誘因和結果上下游關係的綜述。本文就針對睡眠和學習記憶的一篇綜述進行總結Brain neural patterns and the memory function of sleep,以期給讀者一個直觀的認識和回顧。


睡眠對包括記憶在內的良好認知至關重要。常用的記錄睡眠的方法主要有兩種:經皮和顱內電極植入的電生理記錄。根據其電生理特徵,睡眠分爲兩個時相:快速動眼睡眠(REM)和非快速動眼時期(NREM)。用於辨別二者的主要特徵爲:非快速動眼睡眠表現出Sharp-wave ripples (海馬尖波漣漪), Cortical slow oscillations (皮層慢波振盪), δ 波 和Spindles等特徵; REM睡眠主要以θ波振盪爲主。不同特徵的睡眠電特徵精確地反應了潛在神經環路的時相活動。這篇綜述,作者(Gabrielle Girardeau,法國國立衛生研究院、索邦大學的獨立課題組組長)主要回顧了這些電的特徵如何指導我們理解和認識睡眠在記憶鞏固的環路機制中發揮的作用和發生機制。特別關注了海馬θ波振盪和Sharp-wave ripples以及他們是如何協調皮層模式的。最後強調了這些腦電活動模式在保持睡眠內穩態的作用,並展望了睡眠參與記憶相關學習鞏固的未來亟待研究的問題和挑戰。
記憶的形成過程實際上對大腦是個面臨挑戰的過程,大腦要決定哪些新的體驗可以被儲存以及整合到已有的記憶裏,從而得以持續和較正。記憶形成於清醒狀態下,連續的新的感覺體驗。睡眠之後給大腦提供一個機會窗口,即在沒有外部刺激的干擾下,去分類和增強新編碼的記憶。這個過程就叫做記憶的鞏固,進而促進長期記憶的產生或者記憶痕跡(Memory engram,這個過程在清醒時期用以支持信息的再提取)的形成。
在睡眠過程中,參與記憶處理的神經網絡金字塔是被內源性激活的。這些活動可以通過非創傷性表面電極記錄(EEG)或者顱內電極記錄局部場電位(LFP)以及尖峯電位。大量的研究表明這些記錄到的睡眠中的電振盪模式對於理解大腦的工作原理很有幫助,這些電信號包括:振盪(比如θ波),可識別波形的瞬間電位(比如樹突電位),和尖峯電位放電活動類型(上下狀態)。下面就從幾個方面討論這些電信號在睡眠大腦是如何參與學習和記憶中扮演的角色。
非快速動眼睡眠和海馬尖波漣漪(SWR)
海馬具有三個層面的結構,信息流從齒狀回(DG)經過CA3到達CA1,睡眠中重要的一個類型是尖波漣漪複合體 (見圖一的Ripples)。在睡眠中, CA3的錐體神經元自發性興奮,發放同步的簇狀放電。這一活動激活大量的CA1區的錐體神經元。在輻射層,CA3的樹突收到的輸入信息產生尖波。同時在CA1區域,激活的錐體細胞和中間神經元的交互活動會產生快速的振盪事件(200Hz): 漣漪。關於這些電信號參與記憶鞏固的過程,目前存在兩步法理論假說:第一,CA3和CA1的亞細胞羣在某次經歷中已經通過θ振盪協同起來,並形成編碼相關新信息的細胞組合。這些組合的CA3細胞在接下來的睡眠時期就通過SWR期重新激活CA1整合的細胞羣,從而使CA1 和CA3間的連接加強,最終導致記憶的鞏固。和這個理論比較契合地幾個現象包括:在曠場實驗的探索中同步放電的多對CA1錐體細胞會在睡眠SWR時相中保持關聯。在清醒時期活動關聯的持續性在接下來的睡眠中通常被認爲會重現(Replay); 利用大量的方法和手段,類似的現象也存在於共放電模式和位置細胞在清醒中激活,在睡眠SWR中重現。更重要地是,人類也存在類似的現象。
最早爲了研究重現與記憶鞏固的關聯,研究者開發了閉環模式系統 (圖二),干擾睡眠漣漪,發現空間記憶受到明顯損傷。在曠場探索之後,光遺傳學手段在睡眠SWR時相抑制CA1 區的錐體細胞嚴重損害這些細胞集羣的重現。這些結果說明空間記憶重現的缺失是由於缺乏空間地圖的鞏固或者記憶痕跡的形成。其中影響SWR相關的重現有很多因素,新奇刺激就是其中一種。
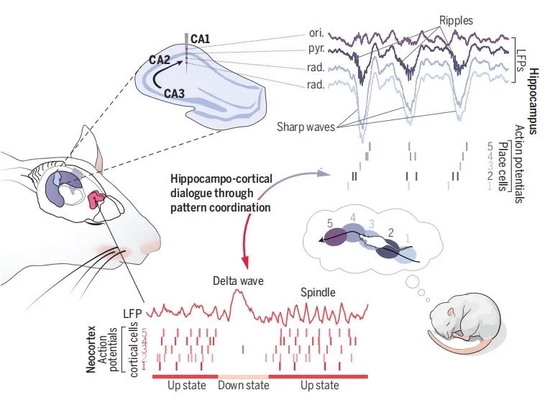
雖然大部分SWR和再激活研究聚焦在CA1區。然而社交記憶痕跡在SWR時相的CA2會再激活,以及CA2的再激活在社交記憶的增強和減弱方面具有雙向調節。這些結果表明,CA3—CA1可能更傾向於SWR再激活的整合,對鞏固空間記憶有幫助。CA2可能對SWR時相在社交記憶方面更傾向。
相對於有限的局部電位漣漪觀察,算法的發展在快速在線監測,特別是在重現內容方面,必將對睡眠重現的理解有極大的幫助。關於這一點的應用,Gridchyn et al。, 訓練了大鼠在兩個不同的環境中覓食,並干擾了接下來的睡眠和靜息態的SWR事件,但不影響在第一個環境中再激活的部分。結果發現大鼠在不被感染的環境中的表現比第二個更好,提示了在第一個環境中的空間記憶的鞏固沒有被打斷從而得以保留。總之,過去幾十年積累的結果提示海馬信息整合的再激活和新鮮信息有關聯。睡眠SWR期的 學習對記憶鞏固很關鍵。但是,令人驚訝的是,海馬的再激活是否也存在於海馬的腹側部分還是未知的。腹側海馬的大腦連接和背側不同,主要與焦慮和應激相關。另外,在非快速動眼期,海馬齒狀回的尖峯放電,反應了強烈的皮層到齒狀回的輸入,被認爲和NREM的鞏固過程有關,但還有待繼續研究。

海馬-皮層的協同通過非快速動眼時期的睡眠模式實現
大部分關於長期記憶鞏固的主要理論包括海馬和新皮層的相互交流。在NREM睡眠期間,皮層環路經歷了高低集羣放電的時相交替。這些改變轉變爲NREM時期局部場電位的經典慢振盪。特別是,下降支和顯著的LFP偏轉有關,叫做δ波。δ波經常伴隨spindles, 來自丘腦的大約10-15赫茲振盪。所有這些皮層節律,大部分通過和其他海馬和皮層模式的相互協調,參與到記憶的鞏固。
人類的經顱刺激可以用於在NREM期間促進慢波振盪。這樣的操作有助於第二天的記憶重獲。大量的和EEG相關的研究表明,慢波和spindle在記憶鞏固中的作用。在齧齒類動物的腦機接口的一個實驗中,非常有趣地發現動物通過訓練可以控制獎賞設備,自我調控預先指定的神經元的放電。在這一操作中,可以觀察到,與任務相關的神經元,在接下來的睡眠事件中,通常會和慢波上升期進行同步化放電。這一實驗進一步證實了慢波振盪和記憶重現地關係。記憶重現的表現基本上符合這樣的規律,即同步化增強,記憶增強。光遺傳地方法去抑制同步化,記憶也會削弱。事實上,目前研究大腦的方法,我們依然侷限於技術,統計和概念上的偏差。我們試圖去研究和觀察我們比較容易去觀察和解析的,比如高羣活動的週期,高放電神經元和顯著的震盪模式。一項原創性研究使用這些方法發現,一些非常稀少的,通常在前額葉被忽略的活動,在明顯的δ波的下降支,實際上在學習過程中是再激活的。
SWR和皮層的NREM睡眠模式在時相上是相互協調的,這一機制被認爲可以促進可塑性和場景(事件)記憶的長期鞏固。海馬SWR的發生頻率在向皮層上下振盪的轉換期和Spindle的波谷增加。用閉環系統來增強海馬和皮層的相關作用,生成一個SWR後的一個spindle複合體下降波,可以增加記憶任務中的表現。光遺傳的方法產生的人工spindle和海馬的慢波漣漪相協調也有增強記憶的作用,突出了“ripple-delta wave-spindle”複合體對鞏固記憶的重要性。而且,海馬慢波漣漪的尖峯部分可以預測接下來δ波的皮層放電,提示了海馬慢波漣漪在皮層信息再激活中的傾向。反過來,皮層放電也可以預測海馬CA1區的信息再激活,睡眠中的感覺刺激可以促進海馬信息的再現和加強記憶,這一現象叫做目標記憶再激活。總結,所有證據表明,記憶的鞏固包含的環路有皮層傾向於記憶信息在海馬的再激活,反過來,海馬的再激活使新皮層的多模型相關信息再激活—這就是海馬和皮層相互協同在記憶鞏固中的基本觀點。
快速動眼睡眠和θ振盪
儘管長期以來,對快速動眼睡眠的主要興趣在於它和人類的生動夢境有關,但是相對於非快速動眼睡眠的研究來講,相關的研究還是缺乏的。REM睡眠的腦電和局部場電位和清醒時的活動非常相似,所以在過去它叫做,“矛盾”的睡眠。實際上,REM睡眠主要是θ振盪,以5-12赫茲爲主。該波在海馬比較顯著,在皮層和皮層下結構也可以記錄到。在清醒期,海馬的θ振盪組織位置細胞放電的有序發放。θ波振盪的精細時間調節,對於空間記憶的編碼和後續的鞏固至關重要,因爲這些過程需要位置細胞在NREM睡眠時相再現。相對來講,很少研究集中在REM睡眠期和θ相關或者不相關的神經元活動。在REM時期,θ波的頻率和幅度的快速增加,成爲相位快速動眼時期,這個時期和整個海馬放電增加和皮層區的協調性有關。相位REM睡眠和來自腦幹的枕骨波有關,被認爲在REM睡眠中協調很多其他結構。
目前未知,REM睡眠θ振盪的特異性變化和行爲學的關聯研究的非常少。然而,海馬的θ振盪和前額葉、杏仁核在厭惡性學習後相互協調增強。利用光遺傳干擾內側隔核來干擾REM睡眠中的θ波會損傷海馬相關的場景相關記憶。另外,海馬齒狀回區成年新生神經元的活動改變,特別在REM時期,可以損傷場景恐懼記憶。雖然該操作並不影響θ振盪,但是無論增強還是減弱放電均會損傷到記憶的鞏固,提示潛在的θ波節律的時間精細調節對新生神經元放電的重要性。有報道也稱,REM睡眠的抑制會引起新生神經元的輕微的突觸結構改變,提示突觸功能減弱。這些結果進一步支持了以往的研究結果,即REM睡眠可以增強新皮層的樹突棘的選擇性增強和減弱。但是更多的工作需要進一步將特定放電的時相精細調節和REM睡眠中θ振盪與已知神經元的樹突棘突觸結構的關係,甚至行爲學輸出之間進行關聯起來,從而真正理解REM睡眠在學習記憶或者其他生理行爲中的功能意義。
拋開海馬皮層談睡眠
根據兩步鞏固法和信息從海馬向皮層逐漸轉移的理論提供的強烈的概念框架,大部分的睡眠模式和記憶鞏固的研究集中在海馬和皮層之間的對話。然而,很多其他結構在記憶的形成中也是參與的。海馬的SWR是非常強大的事件,可以同步化新皮層外的其他結構,潛在的把其他成分中比如情緒基調關聯到不同形式記憶的鞏固中。例如在獎賞體驗後,獎賞定位的海馬位置細胞和獎賞編碼的腹側紋狀體神經元在睡眠的SWR時相和海馬活動同步放電導致紋狀體激活。另外,背腹側海馬調節獎賞處理的另一個重要核團-伏隔核的細胞羣是不同的。在基底外側杏仁核,價值編碼的主要中心,一類亞細胞羣被海馬的SWR所調節。在厭惡空間記憶中,聯合海馬杏仁核的神經元代表區,會在NREM期的SWR時相再現。這些結果提示,海馬的SWR可能是調節整個大腦範圍的調節器,允許跨越皮層的和分佈在皮層外結構的記憶痕跡得以形成。
睡眠節律和可塑性:鞏固和內穩態
學習被認爲與赫布可塑性(Hebbian plasticity)和突觸增強有關。根據突觸穩態假說,睡眠通過下調突觸重量從而防止飽和,進而允許在接下來的清醒時期新記憶的形成,在穩態調節中起着關鍵性作用。特別是,這個模型預示,突觸整體重量在清醒時增加; 在睡眠中減弱。儘管有突觸結構和分子證據支持這一過程,但是在在體和實時衡量突觸結構變化和強度依然很有難度。雖然皮層慢波活動起源於高度同步化的上下狀態,它們的幅度被認爲反應了皮層神經元的突觸強度。相應地,慢波振盪在延長地清醒後最強,然後隨着增加的睡眠事件逐漸減弱。這一現象符合突觸穩態假說。而且,皮層誘發電位的坡度變化-突觸效率的標記之一,和慢波活動的變化相關,提示了慢波可能持續對突觸進行降級。同時,在清晰和睡眠時期的放電動力學用於神經興奮性的代表。和突觸穩態假說模型比較契合的還有海馬細胞,作爲一個羣體,在清醒時期放電是逐漸增加。在睡眠期,整個神經元網絡的活動都是降低的,但是在不同的睡眠時相會有相反的趨勢,比如尖峯活動在NREM期明顯增加,在REM時期減弱。值得注意的是,REM時相放電下調可以通過spindle和SWR發生頻率來預測。最後,經典NREM睡眠的SWR,長期被認爲有助於通過長時程增強促進記憶的鞏固,同時也會觸發長時程抑制。而它們的抑制會阻止睡眠期誘發電位正常減弱,提示了睡眠和突觸可塑性在內穩態中的潛在相互作用。
展望
雖然描述起來簡單,睡眠和記憶之間的關係是個非常複雜的研究領域。首先,睡眠不是同質化的,可以被分爲幾個階段和通過不同的節律和模式分類的亞階段。其次,記憶有很多種(包括事件性、語義性、程序性、技能性和巴普洛夫條件性),這些不同類型的記憶依賴於不同的卻偶有重疊的結構網絡。他們表現出不同的睡眠模式。再者,事件記憶不是完全的真實反映實際事件的。事件記憶的形成,因此,包含信息的最初編碼,修正,和其他記憶整合,甚至遺忘。想象睡眠、記憶和參與結構的複雜性,我們如何設計相關的基礎研究來揭示睡眠在記憶中所扮演的角色是我們面臨的挑戰。
在齧齒類動物身上,NREM睡眠時相傳統地被研究爲一個同質化的時期。怎麼鑑別出NREM特異的亞分期或者微階段,並找到潛在的和人類NREM的三個亞階段相匹配的分期,將是連接它們和各種記憶處理的不同方面到各種行爲學水平的一個切入點。相位性和強直性REM睡眠在人類和其他種屬中都有待研究。同時,研究睡眠的不同模式可能比時期更可靠。閉環系統的發展和腦機接口界在對神經元放電時的實時模式監測,以及EEG或者LFP信號給領域內理解睡眠模式在記憶形成過程中的作用有着非常重要的優勢。雖然有着經驗痕跡的位置細胞序列在接下來的SWR睡眠中會再激活。但是,沒有因果關係的證據表明位置細胞序列本身,對比短時間窗內的位置細胞集羣的少量激活而言,對記憶的鞏固是重要的。檢驗尖峯時間在模式中的重要性理論將需要更精確的實時工具,需要在不干擾特異神經元放電的情況下,干擾在更廣時間維度的時間的精確度。反過來,驗證序列本身的相關性問題潛在的會重新定位領域就目前解碼算法未能識別的近乎80%的SWR相關的神經元內容。這些可能會是我們尚未能夠識別的重激活事件。根據該假設,SWR相關的高同步化事件的主要功能包括那些我們未解碼的,會促進各種記憶播放的鞏固。另外一個新出現的,更綜合的一個理論是:在睡眠中,皮層和海馬由於其生理特性和硬件連接包含的高度同步化活動,進入默認模式。這些模式可能主要用於內穩態平衡的目的。但是清醒活動和記憶編碼可能偏向於精確的放電時間,從而特殊的記憶痕跡從而得到鞏固。而清醒時期的活動會使這種偏向更強,在學習和新奇刺激後更持久,導致SWR時相的重現-噪音比更強。從這個角度,內穩態和鞏固在同一個維度,並強烈的依賴於神經活動在經典睡眠模式的精細時間。
最後,再激活,記憶鞏固的主要機制在腦結構和睡眠時期方面並不普遍適用。目前內穩態在新皮層的研究很多,因此,更多的工作需要被精細的鑑定睡眠模式在非海馬皮層結構參與的記憶處理過程中的作用。同時需要被進一步研究的是,鞏固和內穩態過程在其他結構中不同的或者特別是那些未被檢測到睡眠再激活或放電率分佈不同的結構。這個方向尤其對高度複雜網絡結構比如腦橋、丘腦、下丘腦、籃斑和基質前腦如何在睡眠的不同時期進行控制,以及轉換需要更多的關注和探索。
在日新月異出現的新的記錄方法和分析算法的激勵下,目前大部分研究者正在完成不同記憶類型和睡眠不同階段和亞階段的多維知識空間的機制研究進程中。雖然研究者們可能永遠無法達成一個統一的關於睡眠的記憶功能的理論,但是擴展和精細這些知識空間將允許我們更好的整合鞏固和內穩態,揭示記憶功能在記憶形成所有階段從編碼到重現的記憶鞏固所有過程的的新連接,並連接記憶法和睡眠的其他方面,比如記憶控制,節律和病理等。
原文鏈接:
http://doi.org/10.1126/science.abi8370










